 BMP9 is a new circulating vascular quiescence factor produced by hepatocytes
BMP9 is a new circulating vascular quiescence factor produced by hepatocytes
| |
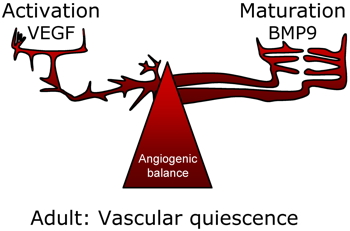
Vascular angiogenic balance (VEGF/BMP9) | |
| |
In 2008, we showed that BMP9 is present in human plasma and serum at biologically active concentrations (5 ng/ml). We further showed that BMP9,
in vivo, inhibited sprouting angiogenesis in the mouse sponge angiogenic assay (David
et al.,
2008)). These results led to propose that BMP9 is a new circulating vascular quiescence factor and that HHT is a BMP disease (see editorial by Bailly S
2008). BMP9 via ALK1 would play a role in the angiogenic balance as a maturation factor against VEGF (vascular endothelial growth factor), which is an activation factor of angiogenesis.