 BMP9 un nouveau facteur circulant de quiescence vasculaire
BMP9 un nouveau facteur circulant de quiescence vasculaire [
en savoir plus]
produit par les hépatocytes [
en savoir plus]
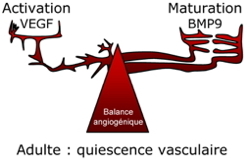
| |
Balance vasculaire VEGF/BMP9 | |
| |
En 2008, nous avons montré que BMP9 est présent dans le sérum et le plasma humain à des concentrations biologiquement actives (5 ng/ml). Nous avons également montré que l'addition de BMP9,
in vivo, inhibe la néo-angiogenèse chez la souris (modèle d'implantation sous-cutanée d'éponge) et entraine une vasoconstriction de la membrane chorio-allantoïdienne du poulet. Ces résultats nous ont permis de proposer BMP9 comme un facteur circulant clé de quiescence vasculaire (David
et al.,
2008) et de proposer un nouveau modèle où la maladie de Rendu-Osler ne serait pas une maladie du TGFß mais des BMPs (voir éditorial Bailly S
2008).